

LOCALISATION OF FUNCTION IN THE CEREBRAL CORTEX

TOPIC CONTENT:
Localisation of function in the brain, motor, somatosensory, visual, auditory and language centres; Broca’s and Wernicke’s areas
Other name for localisation of function in the cerebral cortex are: Cortical Localisation or Cortical Mapping, which specifically refers to identifying the functions of various regions within the cerebral cortex.
DISCLAIMER
Many texts describe this topic as localisation of function in the brain, but this can be misleading. The brain is divided into the hindbrain, midbrain, and forebrain, and these regions handle a variety of autonomic and basic functions. However, when we discuss localisation of function in this context, we are focusing specifically on the Cerebral Cortex, the outer layer of the forebrain responsible for higher cognitive functions like reasoning, language, and voluntary movement. Thus, the primary focus is on the outer surface of the cerebrum—the cerebral cortex.
KEY WORDS FOR: LOCALISATION OF BRAIN FUNCTION
AUDITORY CORTEX: Situated in the temporal lobe, responsible for processing sound information.
CORTEX: General term for the outer layer of the brain's regions involved in processing information.
CEREBRUM: The largest part of the brain, which includes the cerebral cortex and is responsible for higher-order functions.
CEREBRAL CORTEX: The outermost layer of the brain, responsible for higher cognitive functions.
COGNITIVE NEUROLOGIST: A specialist who studies how brain damage or neurological disorders affect cognitive functions like memory, language, and decision-making.
BRAIN LOBES: The four main lobes of the brain, each associated with specific functions:
FRONTAL LOBE: Responsible for reasoning, problem-solving, and motor control.
PARIETAL LOBE: Processes sensory information and spatial awareness.
OCCIPITAL LOBE: Primarily involved in visual processing.
TEMPORAL LOBE: Key for auditory processing and memory functions.
BROCA’S AREA: A region in the frontal lobe associated with speech production.
DISTRIBUTED PROCESSING: The concept that brain functions are not isolated but depend on networks of interconnected regions working together.
EQUIPOTENTIALITY THEORY: This theory suggests that while some basic functions may be localised, higher cognitive functions are more distributed across the brain.
HOMUNCULUS MAN: A visual representation of how different body parts are mapped onto the somatosensory and motor cortices according to the amount of control or sensory input they receive.
LOBES VS CORTICES: Lobes are the broader regions of the brain (e.g. frontal, temporal), while cortices are specialised areas within the lobes that handle specific tasks, such as the visual cortex for vision.
LOCALISATION OF FUNCTION: The theory that certain areas of the brain are specialised for specific functions, such as language or movement.
MOTOR CORTEX: Controls voluntary movements and is located in the frontal lobe.
NEUROIMAGING: Techniques such as fMRI and PET scans that allow scientists to visualise brain activity and better understand the distribution of functions across different brain regions.
PHANTOM LIMB: The phenomenon where individuals who have had a limb amputated continue to feel sensations, including pain, in the missing limb, due to the brain’s sensory map.
PHRENOLOGY: A now-debunked theory that claimed the shape of the skull could determine personality traits and cognitive abilities by mapping bumps on the head.
POST-MORTEM The examination of a body after death to determine the cause of death or study specific conditions, often used in brain research to examine the effects of brain damage on function.
PREFRONTAL CORTEX: The region at the front of the frontal lobe, associated with decision-making, personality, and social behaviour.
SOMATOSENSORY CORTEX: Found in the parietal lobe, it processes sensory inputs from the body, such as touch, pressure, and pain.
TOPOGRAPHICAL MAPPING: Refers to the way the brain organises the body's sensory and motor functions in a map-like representation, as seen in the motor and somatosensory cortices.
VISUAL CORTEX: Located in the occipital lobe, it processes visual information like shape, colour, and motion.
WERNICKE’S AREA: A region in the temporal lobe responsible for language comprehension
Here is the link to BBC Brain Story Documentary with Dr Susan Greenfield, Episode One: "All in the Mind":
Watch BBC Brain Story - Episode 1: "All in the Mind"
LOCALISATION OF FUNCTION IN THE CERBRAL COTEX
INTRODUCTION TO LOCALISATION OF FUNCTION
The cerebral cortex, the outermost layer of the brain, is key in the context of locaisation of brain function. As, unlike other parts of the brain, such as the hindbrain (which regulates autonomic functions like breathing and heart rate and is the same across all humans), the cortex is responsible for higher intellectual abilities, These include reasoning, decision-making, language, and voluntary movement. and could as a result be uniquely organised based on experience..
The cerebral cortex, often referred to as the "wrinkly outer covering" of the cerebrum, is about ¼ inch thick. Not all animals have it—only mammals do—and it was the last part of the brain to evolve. The wrinkles (called gyri and sulci) increase the surface area of the cortex, allowing for more neurons to fit in the brain. This expansion in surface area is linked to higher intelligence. Humans have the most wrinkled cortex, followed by dolphins and primates. These wrinkles enable advanced functions like self-recognition, language, theory of mind, and self-awareness.
Localisation of function in the cerebral cortex refers to the idea that specific regions of the brain, particularly in the cerebral cortex, are responsible for distinct higher brain functions.
Localisation involves identifying and mapping where these functions reside in the cortex, and understanding why certain areas are specialised for particular tasks.
APPLICATION OF LOCALISATION
Localisation serves a practical purpose. It helps scientists understand how brain damage or disease in one area can lead to the loss or impairment of certain functions. For example, damage to Broca’s area, a region associated with language, can result in speech difficulties.
This theory also explores whether functions are consistently located in the same brain regions for all members of a species. By pinpointing specific cognitive tasks to certain brain areas, scientists can better predict how the brain operates and how it responds to injury. This is vital in the treatment of brain injuries, strokes, and neurological disorders.
HISTORY
The theory of localisation of function began in the 19th century, with early neurologists like Marc Dax, Carl Wernicke, and Paul Broca, making significant discoveries. They showed how damage to specific areas of the brain affected particular functions, such as speech and language, sparking the idea that cognitive tasks could be mapped to specific regions.
During this time, phrenology—the belief that traits and behaviours could be mapped to bumps on the skull—gained popularity. Although ultimately discredited, phrenology led to important research. In the 1960s, David Hubel and Torsten Wiesel were initially asked to investigate phrenology, but their work on the visual cortex resulted in groundbreaking discoveries. They demonstrated that certain neurons in the brain were specialised to respond to specific stimuli, such as shapes and edges, further reinforcing the idea of functional specialisation.
EVALUATION AND COMMENTARY
REASONS FOR EVOLUTIONARY ADAPTATION
From an evolutionary point of view, localisation makes evolutionary sense; the development of the human cerebral cortex is believed to have been driven by a variety of selection pressures arising from both the physical and social environment. Key factors such as climate, diet, food availability, sexual selection, group size, coalition formation, and parental care played crucial roles in shaping the brain's evolution.
Evolutionary psychologists propose that the human brain evolved through natural selection, with specific brain regions—such as Broca's area—developing in response to environmental pressures that favoured behaviours increasing chances of survival and reproduction. For instance, behaviours that promoted group coordination, problem-solving, and social bonding would have been favoured over time, leading to the development of specialised brain areas.
Language is thought to have arisen as a direct result of hunting. Humans, lacking the physical attributes typical of successful predators—such as speed, sharp teeth, or a strong sense of smell—relied on group hunting and Machiavellian tactics to survive. To hunt successfully, more sophisticated forms of communication were essential. These would have enhanced group coordination and, in turn, improved survival rates.
In this context, regions involved in complex communication, such as Broca's area, likely evolved to meet both social and survival needs, making the brain more adaptable and sophisticated. The development of the cerebral cortex, therefore, reflects the evolutionary importance of such behaviours in the human species.
INDIVIDUAL DIFFERENCES
One of the main issues with the localisation of function in the cerebral cortex is its limited applicability to the general population. Individual differences, such as the brain’s capacity to adapt and rewire based on environment and upbringing, mean the theory does not universally apply to everyone. Human brains are highly malleable at birth, with neural connections shaped significantly by external factors. Other key individual differences are discussed below
LEFT-HANDERS
For instance, a major challenge to the theory of localisation of function is that not all individuals have the same brain organisation. In particular, about 40% of left-handers have their brain functions arranged differently, with Broca’s area (responsible for speech production) located in the right hemisphere instead of the left, and the visual-spatial processing area situated in the left hemisphere. Even among the 60-70% of left-handers who use the left hemisphere for language, their brain wiring differs from right-handers because their brains must connect and integrate information across both hemispheres more extensively. This is especially true for the small proportion of left-handers with a bilateral language organisation, where both hemispheres are involved in language processing.
As a result, the theory of localisation, which assumes that specific functions are always tied to specific regions, does not hold true for many left-handers and even some right-handers.
MALE VERSUS FEMALE BRAINS
There are notable differences in brain structure between males and females, particularly in language-related areas. Research by G. Pearlson has shown that two key regions in the frontal and temporal lobes—both linked to language processing—are significantly larger in women. This suggests a biological basis for women's superiority in language skills. In the study, magnetic resonance imaging (MRI) was used to measure grey matter volumes in 17 women and 43 men. The results showed that Broca's area was 23% larger in women, while Wernicke's area was 13% larger. These findings were later corroborated by researchers at the School of Communication Disorders at the University of Sydney, Australia. They reported that Wernicke's area was 18% larger in women, while Broca's area was 20% larger.
Additionally, studies have indicated that the corpus callosum—the neural bridge connecting both brain hemispheres—is generally larger in women. However, this finding has been challenged by more recent studies.
These anatomical differences suggest that localisation of function may not apply uniformly across genders. The theory often fails to consider the distinct brain organisation seen in males and females, indicating a level of beta bias, where differences between the sexes are overlooked. Variations in cortical volume and patterns of activation during language tasks highlight that the localisation of function theory might need to be adapted to better reflect male versus female brain structures.
NON-TYPICAL BRAINS
Similarly, people who are congenitally blind or deaf will have differently organised auditory and visual cortices due to their disabilities. In these individuals, primary functions like sight and hearing may not be processed in the same way as in typical brains. For instance, areas of the brain that would normally handle visual or auditory input may be pruned, meaning they shrink or reorganise because they are not receiving stimulation. Instead, these brain regions are often recruited to enhance other intact senses, such as touch or hearing for blind individuals, or vision for those who are deaf. This shows that localisation of function can vary significantly depending on individual experiences
SUMMING UP THE RESEARCH ON LOCALISATION OF BRAIN FUNCTION
Despite individual differences in brain organisation, approximately 90% of people do have localised areas for specific functions, such as language and motor control. Numerous research methods provide strong evidence for this claim.
The problem with some of the earlier research methods was that they were non-experimental, such as case studies, ablations, and post-mortems. For example, participants like Paul Leborgne, famously known as "Tan", may have had individual differences in brain organisation. “Tan” might have had a larger speech area than other people, especially if he had been bilingual, making it difficult to generalise the findings from one individual to the wider population. Additionally, post-mortems only show brain structure after death, providing no insight into real-time brain activity. Similarly, ablations—the surgical removal of brain tissue—are imprecise, as their effects can vary depending on the individual’s brain organisation or even the species being studied, which limits generalisability.
Since the 19th century, more advanced methods have been introduced, such as electrical stimulation, which allows for more precise mapping of smaller brain areas, like the topographical maps in the motor and sensory cortices. These experiments have revealed how motor and sensory functions are systematically organised. However, these methods often use animals, whose motor and sensory systems differ from humans. For instance, animals may have tails, whiskers, or superior hearing and smell, which makes complete generalisation to humans difficult.
Since the 1990s, the localisation of function in the cerebral cortex has been strongly supported by neuroimaging techniques, such as fMRI and PET scans. These methods are far more advanced, as they can be used with thousands of human participants. Unlike older techniques, scans are non-invasive and allow researchers to observe real-time brain activity. This means that scientists can confidently assume that most people have brain functions that are organised or localised in similar areas.
While these neuroimaging studies strongly support the role of Broca’s and Wernicke’s areas, some more recent research has provided contradictory evidence. For example, Dronkers et al. (2007) conducted an MRI scan on Tan’s brain to confirm Broca’s findings. Although they found a lesion in Broca’s area, they also discovered evidence suggesting that other brain areas might have contributed to his speech production difficulties. Additionally, other neuroscientists have found that Broca’s area is active during cognitive tasks unrelated to language. Fedorenko (2012)found that Broca’s area has two regions—one involved in language and the other responding to various demanding cognitive tasks, such as solving maths problems. These findings suggest that Broca’s area may not be the only region responsible for speech production, and the deficits seen in Broca’s aphasia may also result from damage to neighbouring regions.
BETTER ALTERNATIVE THEORIES
Despite advancements in research, strict localisation of function has not been universally accepted. Jerry Fodor’s Modularity of Mind theory offered a more nuanced view, suggesting that the brain is organised into distinct modules—areas specialised for certain cognitive tasks. These modules function independently but must interact to produce complex behaviours. However, Fodor's theory focuses more on cognitive processing rather than neural localisation and is more relevant to understanding how different brain systems work together rather than challenging the physical localisation of functions like motor control or language.
In the 1960s, early cognitive psychologists such as Atkinson and Shiffrin (1968) and Baddeley (1974) introduced memory models that contributed to a more system-oriented view of brain function. Their multi-store models of memory implied that cognitive processes like memory storage and retrieval rely on interconnected systems. While their work did not directly challenge the localisation theory, it suggested that cognitive functions might not be confined to specific brain regions but are distributed across systems that work together. This idea aligns more closely with modern theories of distributed processing than strict localisation.
The theory of distributed processing became more prominent in the 1990s as neuroscience and neuroimaging techniques revealed that tasks such as language, memory, and motor control involve communication between multiple brain regions rather than being confined to a single area. Parallel processing is a key example of this, especially in the visual system, where the brain simultaneously processes different aspects of a visual scene—such as colour, motion, shape, and depth—across multiple regions before integrating this information into a unified perception. This complex interaction demonstrates how brain functions are distributed across networks rather than being strictly localised.
Also relevant to challenging localisation theory is Karl Lashley’s equipotentiality theory, which argued that higher cognitive functions like memory and learning are not confined to specific regions. Lashley found that if one part of the brain was damaged, other parts could often compensate for the lost function, suggesting that the brain has a more distributed and flexible capacity for processing information. This contradicted the idea of strict localisation, particularly for higher cognitive tasks.
DISTRIBUTED PROCESSING
Another theory which suggests multiples regions is Distributed processing processing. This theory also suggests that brain functions are not isolated to single areas but depend on networks of interconnected regions working together. For example, while Broca’s area and Wernicke’s area are essential for language production and comprehension, it is the connection between these regions and others that enables fluent communication. Damage to the pathways linking these areas can result in impairments resembling damage to the areas themselves, reinforcing the idea that multiple regions collaborate to support cognitive functions.
This model highlights that brain function is not just about where specific tasks are located but about how different areas interact. As such, distributed processing presents a more integrated and networked view of brain function, contrasting with the older notion of rigid localisation.
CURRENT THEORY OF BRAIN LOCALISATION WITH RESEARCH
Today, the theory of localisation of brain function continues to be widely accepted but with greater emphasis on the interaction between localised regions and distributed processing. Modern research supports that certain areas of the brain are specialised for specific tasks (e.g., Broca’s area for speech production and Wernicke’s area for language comprehension). However, there is also a growing recognition that complex cognitive functions—such as memory, language, and decision-making—rely on the integration of multiple brain regions rather than being strictly isolated to one.
For example, neuroimaging studies using techniques like fMRI and PET scans have shown that even tasks traditionally thought to be localised (like speech or motor control) activate widespread brain networks. Fedorenko’s (2012) research, which found that Broca’s area has subdivisions that are responsible not just for language but also for more general cognitive tasks, further supports the idea that brain regions may not be as strictly specialised as once thought.
NEUROPLASTICITY

Additionally, research on brain plasticity has shown that the brain can reorganise itself after injury, with undamaged areas compensating for lost functions. Functional plasticity refers to the brain’s remarkable ability to adapt after injury by redistributing functions to undamaged areas. When a specific region of the brain becomes damaged, functions typically associated with that area can be reassigned to other parts of the brain, often in the opposite hemisphere or in regions adjacent to the damaged area.
For example, if the language centre in the left hemisphere is damaged, the right hemisphere or surrounding areas of the brain can sometimes take over language processing. This adaptability casts doubt on the strict localisation of function theory, as it shows that cognitive abilities are not necessarily fixed to one region. The brain's capacity to reorganise itself suggests that functions are more distributed than the localisation theory traditionally implies.
This flexibility demonstrates that while certain functions are localised, the brain operates as an interconnected system, capable of adapting and redistributing tasks when necessary.
LOCALISATION REMAINS USEFUL
Despite these challenges, the theory of localisation of function remains a crucial tool in understanding how the brain works. Knowing which regions are responsible for specific tasks helps clinicians diagnose and treat patients following brain injuries, strokes, or neurological disorders. Localisation of function is especially valuable in treating patients with cognitive or motor impairments, as it allows clinicians to pinpoint the affected brain areas.
CONCLUSION
In conclusion, while the theory of localisation of function has significantly shaped our understanding of brain-behaviour relationships, it’s clear that the brain operates as a more interconnected and flexible system than initially believed. Early localisation theories provided key insights, especially through the discovery of areas like Broca's and Wernicke's regions. However, individual differences in brain organisation, evidence from neuroimaging, and concepts like distributed processing and brain plasticity suggest that cognitive functions are not strictly confined to isolated regions. Future research will likely focus on how localised functions interact within a broader, dynamic neural network, providing a more integrated understanding of human cognition
WHAT HAS BEEN LOCALSIED IN THE CEREBRAL CORTEX
Now that we have examined what localisation of function means in the brain and discussed its importance, it is time to focus on the specific functions that have been discovered and localised. These are primarily concerned with the following key areas:
Motor Centres
Somatosensory Centres
Visual Centres
Auditory Centres
Language Centres: Broca’s and Wernicke’s Areas
Each of these areas plays a vital role in higher cognitive and sensory functions, and their localisation helps us understand the brain's structure and functioning in more detail
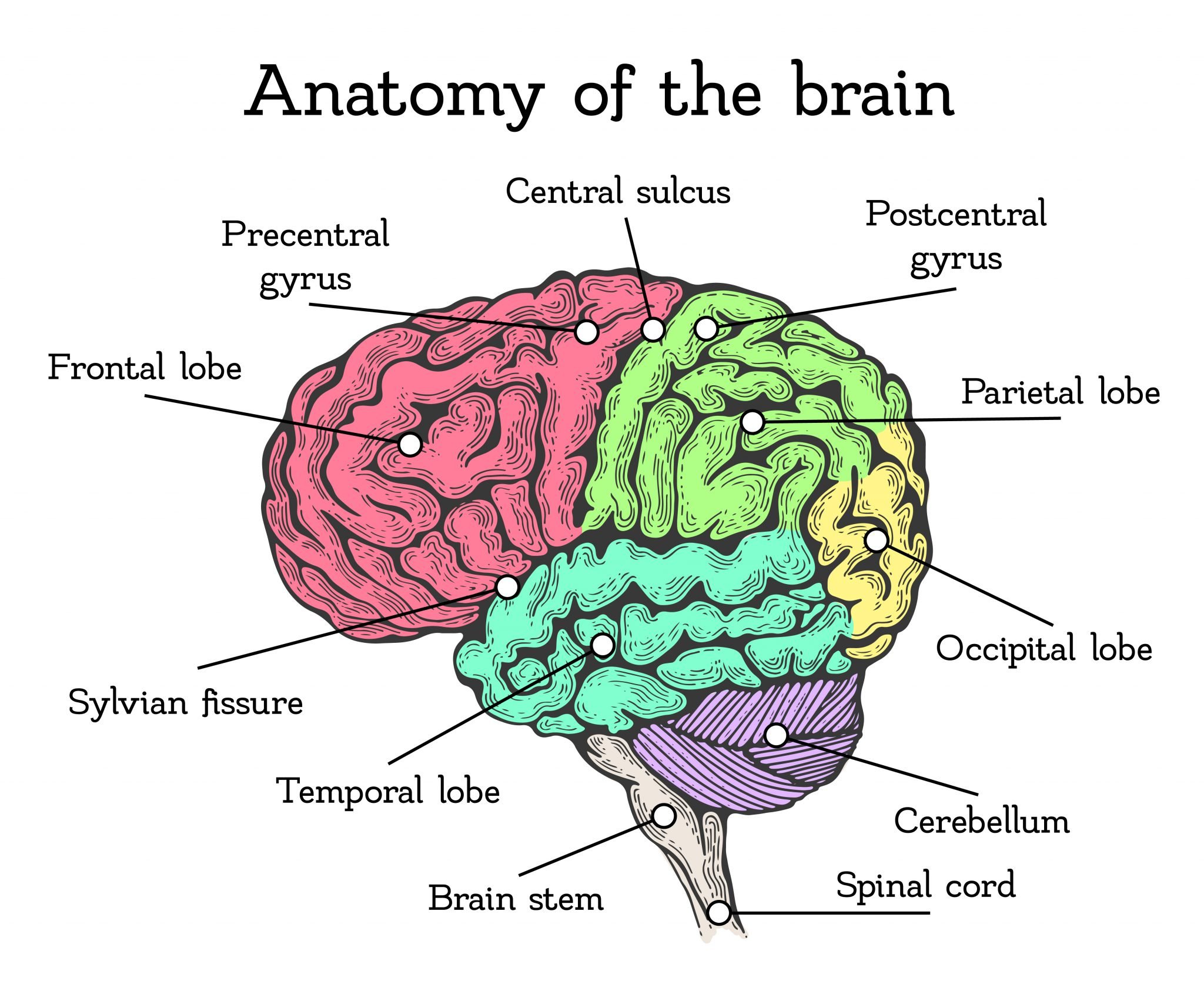
LANGUAGE CENTRES IN THE BRAIN
BROCA’S AREA

BROCA'S AREA AND SPEECH PRODUCTION
Have you ever wondered how you're able to speak, or why your mouth moves in a specific way when forming certain sounds? This all comes down to a region in your brain called Broca’s area. Essentially, this part of the brain is responsible for remembering the motor movements needed for speech production.
For example, when you say the letter "T" in English, your tongue taps your teeth. Similarly, you can’t pronounce the letter "B" without moving your lips. Every time you speak, your tongue, larynx, and pharynx configure in a precise way to make you understood. These movements are learned from a very young age. Think back to when you first struggled to pronounce words, or when babies begin experimenting with sounds like "bbb" or "ddd" as they vocalise consonants in their native language. All these early vocalisations help train Broca’s area to coordinate the intricate muscle movements involved in speaking.
Once these speech motor patterns are established, they become automatic, meaning we don’t consciously think about how to move our mouth to speak. But when Broca’s area is damaged, such as through a stroke or brain injury, a person loses the ability to produce coherent speech. This condition is known as Broca’s aphasia.
In Broca’s aphasia, individuals can often understand language but struggle with speech production. Their speech becomes fragmented, slow, and effortful, as they cannot coordinate the necessary muscle movements to articulate words. For instance, they may know exactly what they want to say but are unable to control their tongue, lips, or vocal cords to form the words correctly. This reveals how essential Broca’s area is in linking thought to speech production, and how its disruption can affect even the most basic forms of verbal communication.
SUMMARY OF BROCA’S AREA
Speech production and articulation
Where is it? The left hemisphere, frontal lobe. The motor region, located in Broca’s area, is close to the area that controls the mouth, tongue and vocal cords.
Research Type: Postmortem, near imaging, split-brain research, electrical stimulation in surgery only ( animals cant speak so no animals reserach here).
Broca's and Wernicke’s area rely on similar research methods, including post-mortems, brain scans, split-brain research, and electrical stimulation during surgery. These methods are crucial for investigating both structural and functional aspects of these language regions.
PLEASE NOTE that research on language areas can only be conducted on humans because we are the only species with fully developed language capabilities. While animals may have forms of communication, they do not possess the complex structures and functions required for language comprehension and production, such as those found in Broca's and Wernicke's areas.
TYPES OF RESEARCH METHOD USED TO LOCATE THE LANGUAGE CORTICES
Research into the localisation of language function has provided a wealth of evidence for distinct areas of the brain, .
POST MORTEMS AND BRAIN SCANS
Post-mortems allow researchers to observe damage in specific brain areas after death, helping identify which regions are associated with language production and comprehension. Brain scans, such as fMRI and PET, offer real-time insights into how these regions activate during language tasks, providing a dynamic view of brain function.
ELECTRICAL STIMULATION IN SURGERY
This method is only possible during human surgeries, as it requires live brain activity for accurate observation. Electrical stimulation cannot be conducted experimentally on humans due to ethical concerns—it is invasive. However, during surgeries to isolate tumours near language areas, electrical stimulation is used to map Broca’s and Wernicke’s areas by blocking action potentials and observing the effects on language production and comprehension. This type of research is crucial because, unlike post-mortem studies, real-time responses are necessary to map the precise areas responsible for language function
RESEARCH FOR BROCA’S AREA:
POST-MORTEM AND EARLY RESEARCH FINDINGS
1825: JEAN-BAPTISTE BOUILLAUD
Jean-Baptiste Bouillaud was one of the first to present cases where frontal lobe lesions led to a loss of speech. Although his work did not fully map specific language centres, it was one of the earliest indicators that language might be localised in certain parts of the brain, particularly the frontal lobes. This early research laid the groundwork for Broca’s later discoveries. His findings align with the idea that speech production is linked to the frontal lobe, supporting the theory of Broca’s area.
1836: MARC DAX
Marc Dax presented cases where damage to the left hemisphere led to speech deficits. His work was significant as he was one of the first to associate hemispheric lateralisation with language, demonstrating that language impairments often occur after damage to the left hemisphere. This observation paved the way for Broca’s later work and strongly supports the theory of language lateralisation in the left hemisphere.
1860: PAUL BROCA
Paul Broca’s research was critical in establishing the localisation of language production. Broca treated a patient known as ‘Tan’, who could only say this one word despite understanding language. Upon performing a post-mortem, Broca found a lesion in the left frontal lobe. Further studies on eight other patients with similar speech deficits and left frontal lobe damage supported his theory that Broca’s area, located in the posterior portion of the left frontal lobe, is crucial for speech production.
However, while Broca’s area has been strongly linked to speech production, recent studies have shown that this region is not exclusive to language tasks. Neuroscientists have found that Broca’s area is also activated during non-linguistic cognitive tasks. For example, Fedorenko (2012) discovered two regions within Broca’s area: one specialised for language and the other involved in general cognitive tasks, such as solving mathematical problems. This suggests that while Broca’s area is crucial for language, it is also part of a broader network for complex cognitive processing.
1864: JOHN HUGHLINGS JACKSON
John Hughlings Jackson contributed to early discussions on loss of speech after brain injury. His observations supported Broca’s work, further establishing that damage to specific brain regions can result in distinct language deficits. Jackson's work reinforced the link between frontal lobe damage and language impairments, aligning with the theory of localised brain function in the left hemisphere.
THE AUDITORY CORTEX
Detection of Sound Quality (Loudness, Tone, Volume, Tempo, Pitch)
The auditory cortex is crucial for analysing and processing acoustic information, dealing with various sound dynamics such as volume, tempo, and pitch. It handles the initial sounds we hear and, through a process called parallel distributed processing, sends different aspects of the sounds to various parts of the brain for further processing—much like an assembly line in a factory. The auditory cortex is bilaterally organised, meaning it exists in both temporal lobesof the brain. This bilateral structure enhances spatial awareness and allows for a more comprehensive interpretation of sound.
While the auditory cortex processes basic sound features like loudness, tone, volume, tempo, and pitch, the content of the sound—what the sound actually is—is processed differently. This type of information is sent to specific areas of the brain for specialised processing, creating some lateralised specialities within the auditory system.
For example, verbal information—such as native languages, second languages, nonsense words, and even backwards speech—is processed predominantly in the left hemisphere. The area responsible for this is known as Wernicke’s area, located primarily in the left temporal lobe. Its function is to handle the semantics of speech, meaning it helps us understand the meaning behind the sounds. If Wernicke’s area is damaged, a person can still hear speech but will struggle to comprehend what is being said, demonstrating that its role is not in interpreting sound but in decoding meaning.
In contrast, non-verbal sounds—such as animal noises, music, and onomatopoeia—tend to be processed with a right-brain advantage. The right hemisphere specialises in interpreting emotional tones in speech (intonation) and in recognising environmental and musical sounds.
This lateralisation likely evolved to aid survival, enabling early humans to distinguish important sounds, such as animal calls or environmental noises, from human speech. This would have helped them differentiate between predator and prey sounds during hunting

RESEARCH ON THE AUDITORY CORTEX
1907 – Pierre Marie and Auguste G. Lhermitte: These researchers first identified the primary auditory cortex in the superior temporal gyrus, specifically in the Heschl’s gyrus of the temporal lobe. This region is essential for processing auditory information such as sound frequency and pitch.
1950s – Wilder Penfield: In his pioneering work on brain stimulation during surgeries, Penfield mapped out the auditory cortex by electrically stimulating different regions of the temporal lobe while patients were awake. The patients reported hearing various sounds when the auditory cortex was stimulated. His work was instrumental in identifying how the brain processes auditory stimuli.
1974 – Michael Merzenich: Merzenich conducted significant research into the plasticity of the auditory cortex. His work with monkeys demonstrated that the auditory cortex could reorganise itself when exposed to new auditory stimuli, suggesting that this region is flexible and can adapt to changes in sound input.
2002 – Sophie Scott and Richard J.S. Wise: In a study using brain imaging techniques, Scott and Wise examined how different parts of the auditory cortex responded to speech and non-speech sounds. Their research confirmed that speech sounds are processed primarily in the left hemisphere, while non-verbal sounds (such as music or environmental sounds) are processed in the right hemisphere. This research provided further evidence of lateralisation in the auditory cortex.
Brain Scans (fMRI and PET): Modern imaging techniques such as functional MRI (fMRI) and positron emission tomography (PET) have shown that the auditory cortex is organised in a tonotopic manner, meaning that different frequencies of sound are processed in distinct areas of the cortex. These scans have demonstrated that while the auditory cortex is primarily involved in detecting sound, it also plays a role in processing complex auditory information, such as music and speech.
Auditory Mapping: Research has demonstrated that the auditory cortex contains a detailed map of sound frequencies, with different areas of the cortex corresponding to different pitches. This tonotopic map is similar in function to the visual map found in the visual cortex and the motor/sensory maps in their respective cortices.
Phantom Sounds and Tinnitus: Research into phantom auditory sensations, such as tinnitus (ringing in the ears), has also contributed to the understanding of the auditory cortex. Studies suggest that tinnitus is caused by maladaptive changes in the auditory cortex, where the brain continues to process sound even in the absence of auditory input. This research highlights the brain’s capacity for reorganisation and the role of the auditory cortex in both normal and abnormal hearing functions.
Brain scans of patients experiencing tinnitus or auditory hallucinations show increased activity in the auditory cortex, even in the absence of external sounds, providing further evidence for the brain’s involvement in processing and misprocessing auditory information.
WERNICKE'S AREA AND LANGUAGE COMPREHENSION

WERNICKE’S AREA: LANGUAGE COMPREHENSION
1874: CARL WERNICKE
In 1874, German neurologist Carl Wernicke identified another key area responsible for language comprehension. His work focused on patients who could speak but were unable to understand spoken or written language, a condition later termed Wernicke’s aphasia. Post-mortem studies revealed that these patients had lesions in the left temporal lobe, specifically in what is now known as Wernicke’s area.
Wernicke’s research proposed that language involves separate motor and sensory regions. The motor aspect, centred in Broca’s area, controls speech production, while the sensory aspect, located in Wernicke’s area, is involved in the understanding of language. Wernicke’s area is positioned close to the auditory and visual regions, making it ideally suited to receive and process sensory input related to language.
While Broca’s area is crucial for producing speech, Wernicke’s area plays an equally important role in understanding language. Located in the left temporal lobe, Wernicke’s area is responsible for processing and interpreting spoken and written language, making it vital for language comprehension.
When you listen to someone speak, your brain doesn’t just hear sounds—it decodes them into meaningful words and sentences. This is Wernicke’s area at work. For example, when you hear the word "apple," you immediately know it refers to a fruit because Wernicke’s area processes the sound and connects it to your mental library of word meanings. This region allows you to understand not only individual words but also complex sentences and nuances in language, such as tone, context, and grammar.
Similar to how Broca’s area is involved in speech production, Wernicke’s area helps you understand language automatically. When functioning correctly, you don’t consciously think about what each word means; the brain processes it seamlessly. However, when Wernicke’s area is damaged, typically due to stroke or injury, a condition known as Wernicke’s aphasia occurs.
People with Wernicke’s aphasia can still produce fluent speech, but the words they use often don’t make sense or fit together logically—this is sometimes referred to as a “word salad.” They may speak in long, flowing sentences but use incorrect words or invent new ones, making their speech difficult to understand. What’s more, individuals with Wernicke’s aphasia struggle to comprehend language, so they might not realise that what they’re saying is incoherent. For example, when asked to name an object like a "pen," they might describe it as something completely unrelated, such as "a green bird," without recognising the error.
In essence, while Broca’s area allows you to articulate your thoughts, Wernicke’s area enables you to understand the language you hear or read. Damage to this region highlights how critical it is for interpreting the world of words around us, turning sound into meaning, and how its impairment can lead to significant communication challenges
SUMMARY OF WERNICKE’S AREA
Language Comprehension and Processing
Where is it?
The left hemisphere, temporal lobe. Wernicke’s area is located near the regions responsible for auditory and visual processing, enabling the understanding of spoken and written language.
Research Type:
Postmortem, neuroimaging, split-brain research, electrical stimulation in surgery (only possible in humans, as animals cannot comprehend or produce human language)

THE LANGUAGE CIRCUIT
When discussing language areas in the brain, it's important to recognise that Broca’s area and Wernicke’s area don’t work in isolation – they form part of a language circuit. These areas are connected by a bundle of nerve fibres called the arcuate fasciculus, and they constantly interact to make speech and understanding possible.
Broca’s area helps us produce speech, while Wernicke’s area helps us understand it. However, they don’t work separately—communication between them is vital. For example, when we listen to someone talking, Wernicke’s are a processes what’s being said, and the information is passed to Broca’s area, which helps us form a verbal response. Without this connection, our ability to have smooth conversations would be seriously affected.
Understanding these areas in isolation is limited because they rely on each other. Damage to the pathway between them can result in conditions like conduction aphasia, where a person might understand speech and speak fluently but struggle to repeat words, highlighting how crucial their interaction is.
In short, focusing only on one area without considering the language circuit doesn't give a full picture. The brain’s ability to process language is a result of multiple areas working together, not just isolated functions
RESEARCH FINDINGS FOR WERNICKE
RESEARCH FINDINGS FOR WERNICKE'S AREA
CARL WERNICKE’S POST-MORTEM DISCOVERIES (1874)
Wernicke's initial discovery came through post-mortem research on patients who had lost their ability to comprehend language. These patients could speak fluently but without making sense, a condition later termed Wernicke’s aphasia. In these cases, Wernicke found lesions in the left superior temporal gyrus of the brain, now known as Wernicke’s area. His research demonstrated that damage to this region impairs the ability to understand both spoken and written language while leaving speech production intact.
MODERN CASE STUDIES AND COGNITIVE NEUROSCIENCE
Advancements in cognitive neuroscience have reinforced Wernicke’s findings. Modern case studies involving brain imaging techniques such as fMRI and PET scans show that Wernicke’s area is heavily involved in the semantic processing of language. These studies demonstrate that Wernicke’s area becomes active when individuals are tasked with understanding complex sentences, suggesting that the region is critical for the integration of linguistic information with auditory and visual inputs.
Example Study: A study by Dronkers et al. (2007) used MRI scans to examine patients with Wernicke’s aphasiaand confirmed that damage to the left superior temporal gyrus resulted in severe comprehension deficits, even though speech production was unaffected. The findings support Wernicke’s original post-mortem conclusions about the role of this brain region in language understanding.
COGNITIVE NEUROLOGY AND LESION STUDIES
Cognitive neurology focuses on studying brain function through clinical cases of brain damage. Lesion studies, where areas of the brain are damaged, provide key insights into how Wernicke’s area functions. When Wernicke’s area is damaged, individuals can produce fluent but meaningless speech and exhibit significant difficulty in understanding language.
Case Study Example: The famous case of patient "EC" (Hickok & Poeppel, 2007) involved damage to Wernicke’s area, resulting in fluent speech without comprehension. This case confirmed the lateralisation of language comprehension in Wernicke’s area and the role of this region in processing semantic meaning.
FUNCTIONAL BRAIN IMAGING: fMRI AND PET SCANS
Recent research using functional magnetic resonance imaging (fMRI) and positron emission tomography (PET)scans has mapped Wernicke’s area and its interactions with other brain regions. These techniques have allowed researchers to observe real-time activity in Wernicke’s area when individuals engage in language comprehension tasks. These studies show that Wernicke’s area is not only active during speech comprehension but also interacts with areas in the frontal lobe, including Broca’s area, as part of a broader language network.
Research Example: A study by Friederici et al. (2011) showed that Wernicke’s area, in combination with Broca’s area, is crucial for understanding syntactic structures in language. Their research demonstrated that damage to this network, particularly in Wernicke’s area, severely disrupts a person’s ability to process grammar and word order.
POST-MORTEM AND LESION RESEARCH
Post-mortem studies continue to be a valuable method for understanding the long-term effects of brain damage, especially in individuals who suffered from Wernicke’s aphasia. These studies often reveal neuronal degeneration in the left temporal lobe, reinforcing Wernicke’s original localisation of language comprehension to this specific region. Furthermore, lesion studies in split-brain patients (who have had the corpus callosum severed) show that the left hemisphere remains the dominant area for processing language comprehension, while the right hemisphere is more involved in prosody and intonation.
THE ROLE OF THE ARCUATE FASCICULUS
Modern research highlights the connection between Wernicke’s area and Broca’s area through the arcuate fasciculus, a bundle of neural fibres that links these two regions. Damage to this connection results in conduction aphasia, where individuals can understand language and speak fluently but are unable to repeat phrases or recall words accurately. This emphasises that language processing involves not just the isolated activity of Wernicke’s area but also its interactions with other regions of the brain.
Research Reference: Hickok and Poeppel (2007) demonstrated that the arcuate fasciculus plays a significant role in linking Wernicke’s and Broca’s areas, enabling the integration of language comprehension and speech production.
These research findings confirm that while Wernicke’s area is central to language comprehension, it functions as part of a broader, interconnected language network involving other regions of the brain.
KEY STUDIES
Carl Wernicke (1874) – Post-mortem Research
Wernicke discovered that lesions in the left superior temporal gyrus led to what is now known as Wernicke’s aphasia, characterised by fluent speech but impaired language comprehension.Dronkers et al. (2007) – MRI Scans
Used MRI to examine patients with Wernicke's aphasia and confirmed damage in the left superior temporal gyrus. This study reinforced Wernicke's findings about the area's role in language comprehension.Friederici et al. (2011) – fMRI Studies
This research used fMRI to show that Wernicke’s area works with Broca’s area to process complex syntactic structures, indicating its role in understanding grammar and word order.Hickok & Poeppel (2007) – Lesion Studies
Explored how damage to Wernicke’s area affects semantic processing. Their work confirmed that lesions in this region result in severe comprehension deficits but intact speech fluency.Poeppel et al. (2012) – Functional Connectivity
This study explored the arcuate fasciculus, a neural pathway connecting Wernicke’s and Broca’s areas, showing how damage to this link causes conduction aphasia, impairing repetition despite fluent speech.Binder et al. (2000) – PET Scans
Used PET to show that Wernicke’s area is active during the processing of semantic meaning, reinforcing its central role in language comprehension
CONCLUSIONS: THE LANGUAGE AND AUIDITOTY CORTICES
HOW THEY WORK TOGETHER
Auditory Cortex: Processes the sound of spoken words, music, and other noises, e.g., their pitch, volume, etc..
Wernicke’s Area: Interprets the meaning of these words (language comprehension).
Broca’s Area: Creates the motor instructions to produce speech (language production).
In short, the auditory cortex handles the primary processing of sound, while Wernicke’s and Broca’s areas handle understanding and producing language. They aren’t subdivisions of the auditory cortex but work together for speech and language-related tasks.
THE MOTOR AND SOMATOSENSORY CORTICES
The motor and somatosensory cortices are two critical areas of the brain that work closely together, despite being located in different lobes. The motor cortex is responsible for voluntary movements, while the somatosensory cortex processes sensory input from the body. Although they have distinct functions, they are anatomically adjacent and share some similarities in how they are organised.
LOCATION AND ORGANISATION
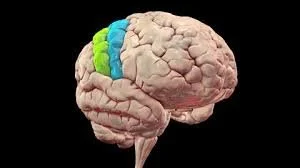
Both the motor and somatosensory cortices are located next to each other, separated by a groove in the brain called the central sulcus. The motor cortex is located in the frontal lobe on the precentral gyrus, while the somatosensory cortex is located in the parietal lobe on the postcentral gyrus.
TOPOGRAPHICAL MAPS IN THE MOTOR AND SOMATOSENSORY CORTEX
The motor cortex and somatosensory cortex are both regions of the brain organised in a topographical manner, meaning that different parts of these cortices correspond to specific areas of the body in a mapped-out fashion. However, the size and placement of these mapped areas are not proportional to the physical size of the body parts they represent. Instead, they are arranged based on the complexity and sensitivity of the movements (in the motor cortex) or sensations (in the somatosensory cortex) they control., reflecting the body's sensory and motor needs rather than its physical layout.
In the motor cortex, which controls voluntary movements, regions responsible for finer, more dexterous movements—like those of the fingers, lips, and tongue—take up disproportionately more space than areas controlling larger, less precise movements, such as those for the back or legs.
Similarly, the somatosensory cortex, which processes sensory information like touch, pressure, and pain, devotes more cortical space to areas with higher sensory input, like the handsand face, compared to the torso or back.

EXPERIMENT: PENCIL TEST
You can test this topographical sensitivity on a willing participant by trying a simple experiment with pencils. If you press two pencils simultaneously on to someone’s back, most people will find it difficult to tell how many points are touching their skin. This is because the back has fewer sensory receptors and takes up less space in the somatosensory cortex. However, if you repeat the experiment on somebody’s hands, they’ll’ll be able to detect both pencil points easily because your hands are more sensitive and have a larger cortical representation.
HOMUNCULUS MAN
The idea of topographical mapping in the motor and somatosensory cortices is often illustrated using the famous "homunculus man." This is a distorted figure where the size of each body part is exaggerated in proportion to how much cortical space is dedicated to it. In the homunculus, the hands, tongue, and face are disproportionately large because these parts of the body require fine motor control or have heightened sensitivity. Meanwhile, body parts like the back or legs, which require less detailed control or sensation, are much smaller in the homunculus representation.
The homunculus gives us a visual way of understanding how the brain prioritises different body parts. For both movement and sensory input, it’s clear that areas requiring more intricate control or detailed sensation are given far more cortical real estate in the motor and somatosensory cortices.

CONCLUSION
The motor and somatosensory cortices are crucial for movement and sensory perception, respectively, and their topographical organisation reflects the brain's priorities. Both cortices dedicate more space to body parts requiring more precision and sensitivity, such as the hands and face, while parts like the back and legs take up less space. The famous homunculus man visually demonstrates these differences, making it easier to understand how the brain maps and processes motor and sensory functions.
THE MOTOR CORTEX IN MORE DETAIL
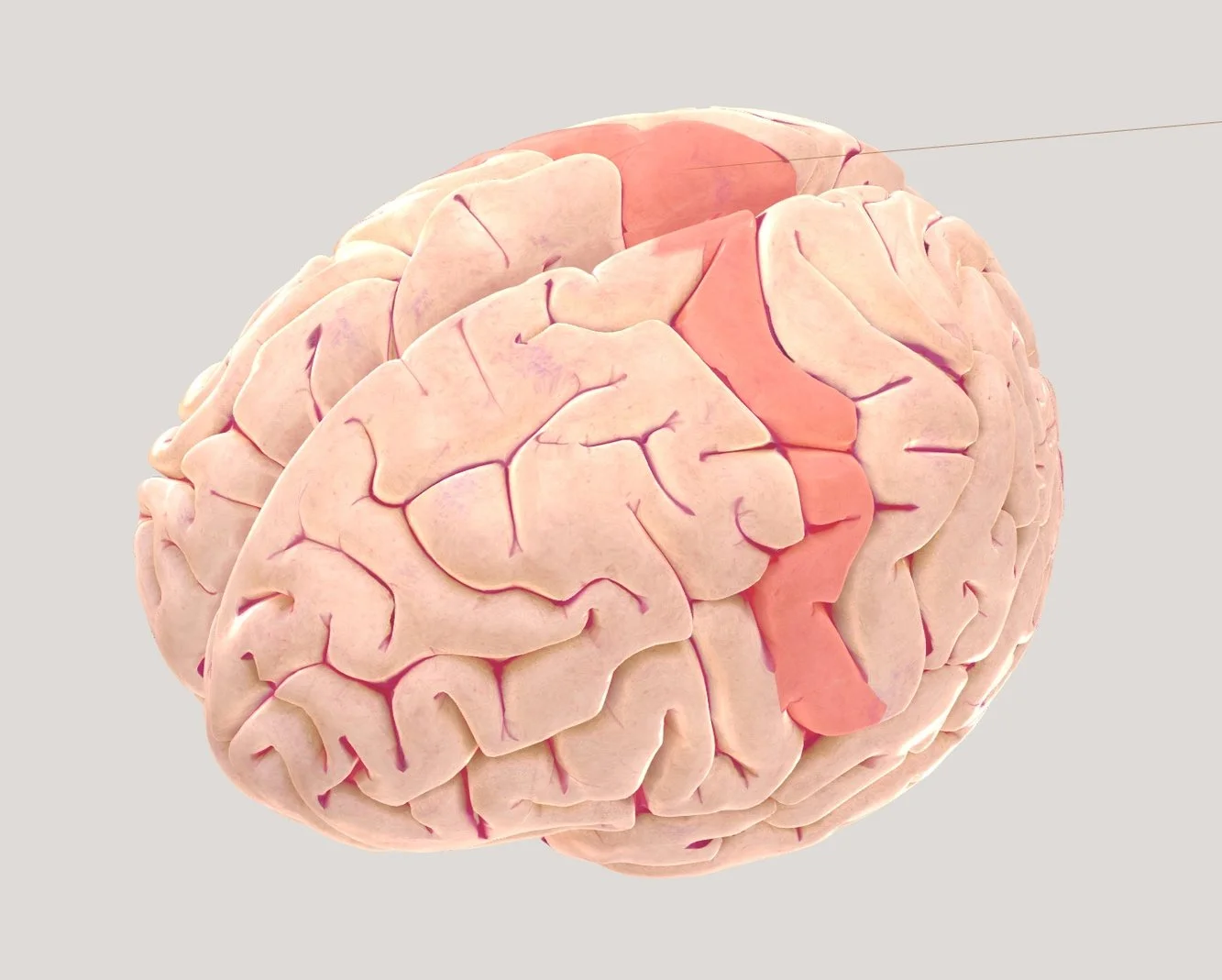
The motor cortex is located in the frontal lobe, along the precentral gyrus, which lies just in front of the central sulcus. This area is responsible for generating voluntary motor movements, controlling muscles across the body. Interestingly, the motor cortex on the right hemisphere controls the left side of the body, and vice versa. The cortex is organised in a way that different regions correspond to specific parts of the body. This organisation is topographical, meaning that areas controlling nearby body parts are located next to each other in the cortex. For example, the region that controls the toes is next to the region that controls the feet, which is next to the region controlling the legs.
Crucially, body parts that require more precise movements, such as the fingers, tongue, and lips, take up more space in the motor cortex than areas like the back or legs, which require less fine motor control. This uneven distribution of space reflects the brain’s need to devote more resources to body parts that perform more delicate movements.
Location: The motor cortex is located in the frontal lobe along the precentral gyrus.
Function: It is responsible for the generation of voluntary motor movements. It exists in both hemispheres: the motor cortex on the right hemisphere controls muscles on the left side of the body and vice versa.
Organisation: Different parts of the motor cortex control different parts of the body. These are arranged logically, so the region that controls the foot is next to the region that controls the leg.
TYPE OF RESEARCH USED TO LOCATE LOCALISATION THE MOTOR, SOMATOSENSORY, AND VISUAL CORTICES
Research into the localisation of the MOTOR, SOMATOSENSORY, and VISUAL cortices has utilised a variety of methods over time, including POST-MORTEM STUDIES, ANIMAL RESEARCH (ABLATIONS AND LESIONS), ELECTRICAL STIMULATION, and NEUROSCIENTIFIC IMAGING TECHNIQUES like fMRI and PET scans. Below is a summary of the research methods used and their relevance to all three cortices.
POST-MORTEM STUDIES AND CASE STUDIES
Historically, post-mortem examinations provided early evidence of localisation by correlating damaged brain areas with specific functional deficits.
MOTOR CORTEX: Damage to the motor cortex (frontal lobe) was linked to paralysis or movement difficulties in the body parts controlled by the affected area.
SOMATOSENSORY CORTEX: Post-mortem studies helped identify that damage to the somatosensory cortex(parietal lobe) resulted in loss of sensation in the corresponding body parts.
VISUAL CORTEX: Damage to the visual cortex (occipital lobe) was linked to blindness or visual impairments, showing the role of this area in processing visual information.
ANIMAL STUDIES (ABLATIONS AND LESIONS)
Ablations (surgical removal of parts of the cortex) and lesions (intentional damage) in animals allowed researchers to study the effects on movement, sensation, and vision.
MOTOR CORTEX: When specific areas of the motor cortex were ablated in animals, the body part corresponding to that area would lose movement. This confirmed the motor cortex's role in voluntary movement.
SOMATOSENSORY CORTEX: Ablations or lesions in the somatosensory cortex caused a loss of tactile sensation, but animals cannot verbalise sensory feedback, limiting the conclusions to behavioural responses like withdrawal from pain.
VISUAL CORTEX: Lesions in the visual cortex of animals caused them to lose their ability to see or interpret visual stimuli, confirming the visual cortex's role in processing visual information.
ELECTRICAL STIMULATION
Electrical stimulation was a breakthrough in localising the motor, somatosensory, and visual cortices, especially during surgeries or experiments with live brain activity.
MOTOR CORTEX: Electrical stimulation of the motor cortex in both humans and animals leads to involuntary movements in corresponding body parts, providing precise localisation of motor control.
SOMATOSENSORY CORTEX: While electrical stimulation can be used on the sensory cortex, it is harder to apply in animals since they cannot verbally communicate sensations. However, in humans, electrical stimulation results in reported sensations like tingling or touch in specific body parts.
VISUAL CORTEX: In humans, electrical stimulation of the visual cortex can result in flashes of light or colour (called phosphenes) being perceived, confirming its role in visual processing. This is more difficult in animals, as they cannot describe what they "see."
PHANTOM LIMB RESEARCH (SOMATOSENSORY CORTEX)
In amputees, phantom limb sensations provide a unique insight into the somatosensory cortex. Even after a limb is removed, individuals continue to feel sensations, including pain, in the missing limb because the brain’s somatosensory map still represents the lost limb.
This research confirms that the somatosensory cortex maintains a topographical map of the body, even when parts of the body are no longer present.
COGNITIVE NEUROSCIENCE AND BRAIN IMAGING (FMRI AND PET SCANS)
Modern techniques like fMRI and PET scans have been critical in real-time observation of the motor, somatosensory, and visual cortices.
MOTOR CORTEX: Scans show which areas of the motor cortex are active when people make voluntary movements, allowing precise mapping of motor functions.
SOMATOSENSORY CORTEX: When people are touched or exposed to different sensory stimuli, fMRI scans reveal activation in the somatosensory cortex, mapping the area responsible for each sensation.
VISUAL CORTEX: Brain imaging also shows activity in the visual cortex when participants are exposed to visual stimuli, confirming its role in processing light, colour, shapes, and motion.
In summary, research on the MOTOR, SOMATOSENSORY, and VISUAL cortices has used a wide range of methods to map their functions. From post-mortem studies and animal ablations to modern neuroimaging, these methods collectively show how each cortex is specialised for different aspects of movement, sensation, and vision. While animal studies laid the foundation, brain imaging and electrical stimulation in humans have provided more precise, real-time evidence of how these areas function.
RESEARCH ON LOCALISATION OF THE MOTOR CORTEX
Research Methods: The localisation of the motor cortex has been studied using a variety of methods including post-mortem analysis, brain scans, split-brain research, electrical stimulation in humans and animals, as well as ablations and lesion studies.
1809: Luigi Rolando first used galvanic current to stimulate the cortical cortex of animals, observing that stimulation in specific areas caused corresponding movements in the animal's body.
1870: Eduard Hitzig and Gustav Fritsch discovered the cortical motor area in dogs using electrical stimulation, providing further evidence of a motor area.
1872: David Ferrier identified specific points in a monkey's cerebral cortex where stimulation produced specific movements. This was a key moment in mapping the motor cortex.
1874: Roberts Bartholow became one of the first to electrically stimulate human cortical tissue to observe motor function.
1875: Sir David Ferrier described various parts of the monkey's motor cortex that control different movements, further refining the map.
1882: Italian neuropsychiatrist Ezio Sciamanna conducted electrical stimulation experiments on a patient with a traumatic brain injury, advancing knowledge of the motor cortex in humans.
1957: W. Penfield and T. Rasmussen developed the motor and sensory homunculus through brain stimulation, providing an accurate representation of human brain function, including the motor and somatosensory areas. Their work is still a foundational reference for understanding cortical maps.
Brain scans and studies confirm that most participants possess a similar motor cortical map, showing the universality of motor function localisation in the brain.
THE SOMATOSENSORY CORTEX IN MORE DETAIL

The somatosensory cortex, located in the parietal lobe along the postcentral gyrus, is primarily responsible for processing sensory information from the body. This includes sensations like touch, temperature, pain, and proprioception (the sense of body position). Like the motor cortex, the somatosensory cortex is organised topographically, with different areas corresponding to different body parts. The left hemisphere processes sensations from the right side of the body, and vice versa.
Just as with the motor cortex, more cortical space is allocated to areas of the body that have a higher density of sensory receptors, such as the hands, lips, and face. This means that you can feel more subtle sensations on your hands or face compared to your back or legsT
RESEARCH ON LOCALISATION OF THE SOMATOSENSORY CORTEX
RESEARCH ON LOCALISATION OF THE SOMATOSENSORY CORTEX
1871 - Silas Weir Mitchell: Mitchell coined the term "phantom limb syndrome" and provided the first detailed account of the condition. Phantom limb syndrome is the phenomenon in which amputees continue to feel sensations, including pain, in a missing limb. This research laid the groundwork for understanding the role of the somatosensory cortex in sensory perception.
1909 - Harvey Cushing: Cushing was the first to electrically stimulate the human sensory cortex. He performed cortical stimulation on 41 patients to map out the motor and sensory cortices during surgery for tumours, trauma, and epilepsy. This allowed Cushing to define the primary motor and sensory cortices and provided critical information about the location and function of these areas.
1957 - W. Penfield and T. Rasmussen: Penfield and Rasmussen created the motor and sensory homunculus, a detailed map of the human cortex that represents the body based on the amount of cortical space each part occupies. Their work revealed how the somatosensory cortex is arranged, with larger areas dedicated to parts of the body with more sensory receptors, such as the hands and lips.
PHANTOM LIMB PHENOMENON: A phantom limb is the sensation that an amputated or missing limb is still present. Approximately 80 to 100% of amputees report sensations in their missing limb, although only a small percentage experience painful phantom limb sensations. These sensations typically resolve within two to three years without treatment. Early theories attributed this phenomenon to a pinched nerve, but Melzack's work demonstrated that the somatosensory cortex plays a central role. Melzack proposed that the body's experience is created by a wide network of interconnecting neural structures within the somatosensory cortex, rather than from a simple nerve injury.
1991 - Pons and Colleagues: Pons and his team at the National Institutes of Health (NIH) showed that the primary somatosensory cortex in macaque monkeys undergoes significant reorganisation following the loss of sensory input. In other words, when the somatosensory cortex no longer receives input from a particular body part (e.g., fingers), the area of the cortex responsible for that part is pruned and reorganised to serve other functions.
Human Evidence: Similar findings have been observed in human amputees. In case studies, amputees who lost a hand showed that the cortical map for the hand reorganised itself after a few months. The cortical area previously devoted to the hand shifted to nearby areas on the somatosensory cortex, such as the face. For example, if someone touched the face of an amputee, the amputee might feel the sensation in their missing hand.
Vilayanur S. Ramachandran: Ramachandran provided further evidence for this reorganisation by showing that stroking different parts of the face in amputees led to the sensation of touch in the missing limb. He hypothesised that phantom limb sensations arise due to this cortical reorganisation in the brain’s somatosensory cortex. Since the brain no longer receives sensory input from the missing limb, it repurposes the neighbouring cortical area (in this case, the face) to maintain a representation of the lost limb.
Brain Scans: Later studies using brain scans on amputees confirmed that cortical reorganisation takes place in the somatosensory cortex, supporting Ramachandran's hypothesis. These scans showed that most people have a clearly defined somatosensory cortical map.
Animal Research: Research on the somatosensory cortex is difficult to conduct in animals, especially for phenomena like phantom limb syndrome, because animals cannot communicate subjective experiences like pain or touch. However, studies on sensory loss in animals, such as numbing parts of their bodies or inducing lesions in sensory areas, provide some insights into how the somatosensory cortex reorganises after sensory input is lost.
The focus on phantom limb research in humans is a powerful method for investigating the localisation and plasticity of the somatosensory cortex, as it highlights how the brain adapts to changes in sensory input.
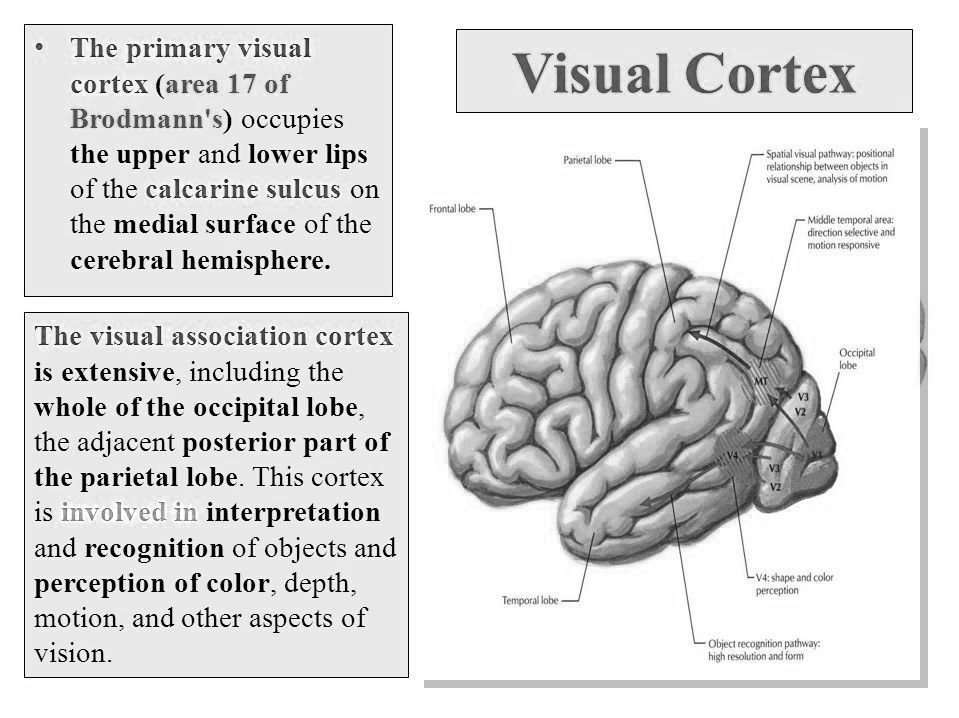
THE VISUAL CORTEX

The visual cortex is located at the back of the brain in the occipital lobe and is responsible for receiving and processing visual information. It plays a crucial role in interpreting everything we see, from colours and shapes to movement and depth. One interesting feature of the visual cortex is how it processes information: visual input from the right-hand side of the visual field is processed in the left hemisphere of the brain, while input from the left-hand side of the visual fieldis processed in the right hemisphere. This cross-wiring helps the brain create a unified picture of the world.
The visual cortex is divided into different areas that specialise in processing specific types of visual information. For example, one area is responsible for processing colour, another handles shape, and yet another processes movement. These specialised areas work together to allow us to understand complex visual scenes
RESEARCH ON LOCALISATION OF THE VISUAL CORTEX

1855 - Bartolomeo Panizza: Panizza was the first to demonstrate that the occipital lobe is essential for vision. His research showed that damage to this part of the brain led to blindness, establishing the occipital lobe as the primary region responsible for visual processing.
1881 - Hermann Munk: Munk provided further evidence by performing ablations (surgical removal of tissue) on the occipital lobe of dogs, which resulted in visual abnormalities. His findings helped to confirm the role of the occipital lobe in visual function and understanding how different regions of the brain contribute to sight.
1947 - Joachim Bodamer: Bodamer, a German neurologist, coined the term "prosopagnosia," also known as face blindness, to describe patients who could no longer recognise familiar faces after damage to the visual cortex, particularly the fusiform face area (a region involved in facial recognition). This condition is one of the key pieces of evidence for the localisation of visual processing in specific areas of the brain.
1981 - David Hubel and Torsten Wiesel: Hubel and Wiesel were awarded the Nobel Prize for their groundbreaking work on the visual system. They mapped out how neurons in the visual cortex respond to different stimuli, such as lines, shapes, and movement. Their work revolutionised our understanding of how visual information is processed and provided critical evidence for the localisation of visual functions in the occipital lobe.
Brain Scans: Modern brain scans have shown that the majority of people have a defined visual cortical map in the occipital lobe. This map represents how the brain organises and processes visual information, including spatial awareness, depth perception, and motion. These scans provide real-time evidence of how the visual cortex functions and confirm earlier anatomical and experimental research
EVALUATION OF RESEARCH METHODS USED IN LOCALISATION OF FUNCTION
One of the main issues with the research into LOCALISATION OF BRAIN FUNCTION is the invasive nature of many methods, especially those involving animals and humans. Techniques such as ABLATIONS (surgically removing brain tissue), LESIONS, and ELECTRICAL STIMULATION have been crucial in identifying specific areas of the brain responsible for certain functions. However, there are several limitations and ethical concerns to consider.
ANIMAL RESEARCH LIMITATIONS
A significant portion of research on brain localisation has involved animals, particularly dogs, monkeys, and rodents. While animals share similar brain structures with humans due to evolutionary continuity, there are substantial differences that limit the generalisability of these findings. For instance, animals have whiskers, tails, and superior senses of smell and hearing. These differences mean that certain areas of their brain, such as the somatosensory cortex, are devoted to whisker movements or tail control, which humans do not possess. Additionally, animals do not process complex languageor recognise faces as humans do. These unique human abilities mean that direct comparisons are not always appropriate. However, these limitations are balanced by the fact that basic neural functions like movement, vision, and sensory processing share evolutionary roots, making animal research relevant.
POST-MORTEM EXAMINATIONS
Many early findings on localisation of function, including the work of Broca and Wernicke, came from post-mortemexaminations. While these have been instrumental in identifying brain regions linked to specific functions, there are several significant drawbacks. Post-mortem studies cannot capture real-time brain activity, making it impossible to observe how the brain functions during cognitive tasks. Furthermore, because they are conducted after death, there is no opportunity to measure plasticity or how other areas of the brain compensate for damage. Additionally, individual differences such as bilingualism—which can lead to different development in Broca’s area—cannot be accounted for, making it difficult to generalise findings to the wider population.
NEUROIMAGING TECHNIQUES
In more recent research, methods like fMRI, PET SCANS, and EEG have revolutionised our understanding of localisation of function by allowing scientists to observe brain activity in real-time without invasive procedures. However, even these methods have limitations. For instance, fMRI can show which areas of the brain are active during specific tasks, but it cannot establish causality—just because a region is active doesn't mean it is solely responsible for the behaviour being studied. Neuroimaging often reveals distributed networks of activity, showing that multiple areas are involved in performing even simple tasks. This challenges strict localisation theories, which suggest that each brain function is housed in a specific location.
ETHICAL ISSUES
The use of invasive techniques, particularly in animals, raises ethical concerns. While these studies have provided valuable insights, the cruelty involved—especially in cases where animals are exposed to pain or distress—cannot be overlooked. As alternatives to animal testing, neuroimaging and TRANSCRANIAL MAGNETIC STIMULATION (TMS) offer less invasive ways to explore brain function in humans. However, TMS is limited in its scope and cannot penetrate deep brain structures as effectively as other methods.
HOLISTIC VIEW OF BRAIN FUNCTION
It’s also important to note that none of these brain areas work in isolation. Research has shown that areas like Broca’sand Wernicke’s areas are part of larger distributed networks that involve other regions of the brain. Modern neuroscience recognises that the brain is an interconnected system, with different regions cooperating to produce complex behaviours. As Marx might suggest, these areas could be seen as the "methods of production", working together to fulfil a common function rather than operating independently.
In conclusion, while research into localisation has provided valuable insights, it is essential to consider the limitations of the methods used. The invasive nature of some techniques, differences between humans and animals, and the growing recognition of the networked nature of brain function all indicate that localisation is not as clear-cut as once believed. Instead, it reflects a more distributed and interactive system within the brain
FOR OTHER EVALATION AND COMMENTARY, PLEASE SEE THE ESSAYS ABOVE AND BELOW
ESSAY: DISCUSS LOCALISATION OF FUNCTION IN THE CEREBRAL CORTEX
MARKS = 16 Marks
A01 = 6 (Outline and description)
A03 = 10 (Evaluate)
SPECIFICATION SAYS:
Localisation of function in the brain: motor, somatosensory, visual, auditory and language centres; Broca’s and Wernicke’s areas.
COMMAND WORDS:
"Discuss" = A01 & A03, i.e., outline and evaluate
STEP ONE: EXPLAIN THE ESSAY TITLE
A01 is an account, outline, or description of a theory or research (APFC: Aim, Procedure, Findings, Conclusion). You should report the essence of a theory or study using appropriate terminology. No evaluation goes here.
The cerebrum is the largest part of the brain, and the cerebral cortex is the ¼-inch thick outer layer of the cerebrum. It is the centre of all higher cognitive functions.
Localisation of function refers to the concept of pinpointing or locating specific cognitive functions, such as language, voluntary movement, and sensation, within the cerebral cortex. This theory asks whether these functions are always located in the same place across individuals and whether damage to a particular area results in the permanent loss of a function.
STEP TWO: DESCRIBE A LOCALISED AREA
Pick 2-3 from the following list as detailed in the specification. While the specification lists five areas of localisation, you are not required to discuss all of them. Depth (focusing on 1-2 theories) is preferred over breadth (covering 3-4 theories) in the A01 section.
Motor cortex
Somatosensory cortex
Language cortices (Broca and Wernicke's areas)
Visual cortex
Auditory cortex
FOR EACH AREA, YOU NEED TO KNOW
What is the function for?
What kind of research supports this? (Post-mortem studies, scans, split-brain research, electrical stimulation in surgery, etc.)
Where is the function located in the cortex?
A01: DESCRIPTION OF THEORY
The cerebrum is the largest part of the brain, and its outer layer, the cerebral cortex, is approximately ¼-inch thick. This layer is responsible for all higher intellectual abilities in mammals, such as reasoning, language, and decision-making. The theory of localisation of function refers to the identification and mapping of these higher cognitive functions—like language, voluntary movement, and physical sensation—to specific regions of the cerebral cortex. It also examines whether these functions are consistently located in the same brain areas across individuals and how damage to these areas might affect functioning.
One key area that has been localised is language. In the 1800s, Paul Broca discovered a region responsible for speech production. He made this finding through a post-mortem on a patient nicknamed "Tan," who had lost the ability to speak except for that one word. Broca found damage to the left frontal lobe of the cerebral cortex, later called Broca’s area, linking this region to speech production.
Similarly, Carl Wernicke identified a region responsible for language comprehension. This area, located in the left temporal lobe, was discovered through post-mortems on patients who could not understand spoken language. The area, now known as Wernicke’s area, is critical for understanding language.
Other functions, such as sensory and motor control, have also been localised. The somatosensory (SS) cortex, situated along the front strip of the parietal lobe, processes sensory and tactile information. It is organised topographically, meaning each part of the body corresponds to a specific location on the cortex, with the left hemisphere controlling the right side of the body and vice versa.
The motor cortex lies adjacent to the somatosensory cortex along the back strip of the frontal lobe and is responsible for voluntary movement. This area has been studied using electrical stimulation, which replicates brain signals, and ablations, where portions of the cortex are surgically removed to observe the effects. Like the somatosensory cortex, the motor cortex is systematically arranged, with more space dedicated to body parts requiring fine motor control, such as the hands and face
EVALUATION OF THEORIES (A03)
Now you can move on to evaluating these points in terms of:
Evidence from case studies and research supporting localisation (e.g., Broca and Wernicke’s work, fMRI scans, and electrical stimulation studies).
Criticisms of localisation, such as Karl Lashley’s equipotentiality theory, which suggests that higher cognitive functions are distributed across the brain rather than localised.
The usefulness of localisation in clinical settings (e.g., treating stroke patients), but acknowledging that distributed processing and brain plasticity may challenge strict localisation.
Discuss the methodological limitations of research methods, such as using post-mortems or animal studies, which may limit the generalisability to live human behaviour.
You could also mention split-brain research and how it has informed localisation theory, especially with regard to hemispheric specialisation
A03 EVALUATION OF THEORY AND RESERACH AND COMMENTARY
Identify your arguments and points.
Evaluate research
Generalisability: Left-handers, non-typical Brains Gender, Accidents/ Plasticity
Distributed parallel functions versus localisation.
Evolutionary adaptive reason for localised areas.
Applied to the real world, e.g., knowing where brain functions are in the cerebral cortex is helpful because …
Any issues or debates
HOW TO AVOID SHOPPING LIST POINTS
For each of these examples, you must avoid just stating a general critique without:
Explaining it fully (what does it mean?),
Applying it to the specific study or theory being discussed,
Evaluating its impact on the findings or conclusions of the research, and
Linking it back to the question to show how it strengthens or weakens the argument.
By developing your points through point-evidence-explain-evaluate (PEEE) or similar frameworks, you will show deeper understanding and avoid the trap of making unconnected, hit-and-miss claims
IDENTIFY THE PEEL POINTS BELOW:
The problems with using methods such as ablations and post-mortems is that participants such as Paul Leborgne aka “Tan” may have individual differences in brain organisation, for example “Tan” may have a larger speech area than other people, especially if the person was bilingual for example. It may, therefore, not be possible to generalise the findings from Tan to other people. Moreover, post-mortems do not show real-time activity in the brain as the person is dead. Similarly, ablations are not a precise research tool, cuts may have different consequences on different brain organisations. However, since this time much more advanced methods of investigating the brain have been introduced, for example electrical stimulation, this method is more precise as it can pinpoint smaller detail such as the topographical maps in the motor and sensory cortices. However, as animals are its main targets, it’s also difficult to generalise as animals have different motor systems, e.g., tails. Lastly, the introduction of scans is the most robust evidence of localisation because …. Scans can use human participants, 1000s of participants can be recruited, it’s not invasive, scans look at real-time brain activity (not dead brains)
This means that researchers can confidently assume most people have functions organised or localised in the same places.
EVALUATING LOCALISATION OF FUNCTION
SUMMING UP THE RESEARCH ON LOCALISATION OF FUNCTION
One limitation of the research into localisation of brain function is the reliance on case studies, ablations, and post-mortem examinations. Case studies, such as the famous example of Paul Leborgne, also known as "Tan", highlight individual differences in brain organisation. For example, "Tan" may have had a larger speech area than other individuals, particularly if he had been bilingual before his injury. This makes it difficult to generalise findings from one person to the broader population. Moreover, post-mortem studies only show brain structure after death and do not provide insights into real-time brain activity, limiting the applicability of their findings. Similarly, ablations (surgical removal of brain tissue) are imprecise, as their effects may vary depending on individual brain organisation or even across different species, reducing the validity of these methods for studying human brains.
On the other hand, since the 19th century, more advanced techniques have been developed, such as electrical stimulation, which allows researchers to map smaller brain regions in more detail. This method can identify topographical maps in the motor and sensory cortices. However, animal studies using these methods face challenges in generalising findings to humans, as animals have different motor and sensory systems (e.g., tails, whiskers) that do not directly translate to human experiences.
In contrast, since the 1990s, neuroimaging techniques such as fMRI and PET scans have offered robust support for the localisation of function in the cerebral cortex. These techniques can involve thousands of participants and are non-invasive, allowing for the observation of real-time brain activity. Additionally, case studies of patients with damage to Broca’s or Wernicke’s areas consistently show that these regions are crucial for language production and comprehension. For example, Broca’s aphasia, caused by damage to Broca’s area, leads to an impaired ability to produce language, while Wernicke’s aphasia, caused by damage to Wernicke’s area, results in difficulties with language perception. The consistency of these findings supports the theory that brain functions are typically organised in the same areas for most people.
LEFT-HANDERS
A significant challenge to the theory of localisation of function is that not all individuals have the same brain organisation. For example, around 40% of left-handers have brain functions that are organised differently, with Broca’s area (responsible for speech production) located in the right hemisphere instead of the left, and the visual-spatial processing area situated in the left hemisphere. Furthermore, even among the 60-70% of left-handers who use the left hemisphere for language, their brains are wired differently from right-handers because they must integrate information across both hemispheres more extensively. Additionally, a small proportion of left-handers have a bilateral language organisation, where both hemispheres are involved in language processing. This variation in brain organisation means that the theory of localisation, which assumes functions are always tied to specific regions, does not apply uniformly to left-handers and, in some cases, even to right-handers.
NON-TYPICAL BRAINS
Similarly, individuals who are congenitally blind or deaf may develop different brain organisation, particularly in the auditory and visual cortices. For these individuals, the areas of the brain that would typically process auditory or visual information may be "pruned" (shrunk or reorganised) due to a lack of stimulation. Instead, these regions may be recruited to support other intact senses, such as touch for blind individuals or vision for deaf individuals. This demonstrates how localisation of function can vary significantly depending on sensory experience.
MALE VERSUS FEMALE BRAINS
There is also evidence suggesting that male and female brains differ in brain structures related to language processing. For example, research by Pearlson et al. has shown that two language-related areas in the frontal and temporal lobes are significantly larger in women. In their study, MRI scans of 17 women and 43 men revealed that Broca's area was 23% larger in women, and Wernicke's area was 13% larger. These findings were later supported by additional research from the University of Sydney, which reported that Wernicke’s area was 18% larger in women, and Broca’s area was 20% larger than in men. Additional studies have suggested that the corpus callosum, which connects both hemispheres, is generally larger in women, although this finding is debated. These anatomical differences indicate that localisation of function theories may not be universally applicable across genders. In fact, they may exhibit beta bias, as they often ignore the structural differences between male and female brains and fail to consider variations in cortical activation during language tasks.
PLASTICITY
Another factor that challenges the theory of localisation is the brain’s ability to adapt, known as functional plasticity. This refers to the brain's capacity to reassign functions from damaged areas to other undamaged regions. For example, if a person sustains damage to their language centre in the left hemisphere, the right hemisphere or other adjacent areas can take over language processing functions. This capacity for reorganisation casts doubt on the idea that cognitive functions are strictly localised in specific brain regions, as other parts of the brain can compensate for damaged areas.
ALTERNATIVE THEORIES
The theory that functions are strictly localised to particular brain areas has been questioned by some psychologists. Karl Lashley, for example, proposed the equipotentiality theory, which argues that while basic motor and sensory functions may be localised, higher mental functions are not. Lashley’s work showed that when one part of the brain is damaged, other areas can often compensate for the lost function, suggesting a more flexible brain organisation.
Wernicke also argued that while different areas of the brain are specialised, they must communicate with each other to produce complex behaviours. One example is a man who lost the ability to read after sustaining damage to the connection between his visual cortex and Wernicke’s area. This suggests that connections between brain regions are crucial for complex tasks such as language, and damage to these pathways can lead to impairments that mimic damage to localised brain regions.
This perspective aligns with the distributed processing model, which emphasises that brain functions are not confined to single regions but are instead dependent on the interaction between multiple brain areas. Critics of strict localisation argue that the brain’s interconnectedness means that understanding how different regions communicate is just as important as identifying where functions are localised.
REASONS FOR EVOLUTIONARY ADAPTATION
From an evolutionary perspective, localisation of function makes sense as the cerebral cortex likely evolved in response to selection pressures from the environment. Factors such as climate, diet, food availability, group size, and social interactions likely shaped the brain's development. For example, the development of language may have been driven by the need for group hunting. Humans, lacking physical advantages like speed or sharp teeth, relied on cooperative hunting and communication to survive. This need for complex communication likely led to the evolution of specialised brain regions such as Broca’s area. Evolutionary psychologists argue that natural selection favoured the development of these specialised areas to enhance group coordination, problem-solving, and social bonding, which in turn improved survival and reproductive success.
APPLICATION TO THE REAL WORLD
The theory of localisation of function has important real-world applications, particularly in the field of psychiatry. For example, future developments in "modular psychiatry" may rely on a modular understanding of the brain. Advanced neuroimaging techniques, which allow for detailed brain mapping, could enable more accurate diagnoses of mental and emotional disorders. Schizophrenia, for instance, has been linked to problems with Broca’s area, demonstrating how understanding brain regions can aid in diagnosing and treating cognitive disorders.
CRITICISMS
Despite the historical importance of Broca’s and Wernicke’s areas, more recent research has challenged the strict localisation of these functions. Dronkers et al. (2007) conducted an MRI scan on Tan’s brain to confirm Broca’s findings. Although they found a lesion in Broca’s area, they also found evidence that other brain areas may have contributed to Tan’s speech production difficulties. Furthermore, research by Fedorenko (2012) has shown that Broca’s area is active during cognitive tasks unrelated to language. She discovered that Broca’s area has two distinct regions: one involved in language, and another responsible for demanding cognitive tasks, such as solving mathematical problems. This suggests that Broca’s area may not be solely responsible for speech production, and deficits in Broca’s aphasia could be due to damage in neighbouring brain regions
A SHORT EXAMPLE OF OTHER POSSIBLE WAYS TO WRITE A01 and A03.
Instead of separating A01 and A03, you can intersperse them throughout the explanation. For example:
A01 general theory of localisation descriptor:
Localisation of function refers to the concept that specific functions, such as language and memory, are localised in certain areas of the brain. EXAMPLE: MPTOR CORTEX One key area believed to be localised is voluntary movement. As early as the 1700s, physiologists began identifying what would later be known as the motor cortex through a method called ablation, where cortical tissue was removed from animals to observe the effects. For instance, when researchers ablated the back strip of the frontal lobe, they consistently observed a loss of motor function in the animals, suggesting that this region was responsible for movement.
A03: However, the use of ablation has limitations. It is invasive and often conducted on animals, making it difficult to generalise findings to humans. Further research refined the technique by removing smaller areas of the brain, allowing scientists to map how the cortex represented the body, resulting in the discovery of the topographical map. This map shows that areas like the fingers and tongue, which require more dexterity, have more cortical tissue devoted to them. This mapping of the motor cortex provides strong evidence for localisation.
A03: More precise methods, such as electrical stimulation, have since been used in human patients, particularly during neurosurgery. For example, when removing tumours, surgeons stimulate the brain to identify essential areas, such as the somatosensory cortex, to avoid damaging them. Research using this method has shown that stimulating the anterior part of the parietal lobe can cause patients to feel sensations in different parts of their bodies, further demonstrating the localisation of tactile processing.
A03: Moreover, phantom limb syndrome in amputees offers additional evidence for the somatosensory cortex. Even after the loss of a limb, patients still feel sensations in the missing limb because the brain continues to represent the limb in the topographical map. This ongoing representation supports the idea that the somatosensory cortex is localised, as it continues to function even when the physical limb is no longer present.
This approach allows for the explanation of key terms (A01) and the inclusion of research and evaluation (A03) in a more integrated way
FURTHER READING
"The Cognitive Neurosciences" (5th edition) – Gazzaniga, M.S. (2019)
Comprehensive exploration of brain function, including localisation, distributed processing, and neuroplasticity.
"Principles of Neural Science" (6th edition) – Kandel, E.R., Schwartz, J.H., Jessell, T.M., Siegelbaum, S.A., & Hudspeth, A.J. (2021)
A detailed examination of neural pathways and brain organisation, covering recent findings on brain function and localisation.
"Human Brain Function" (2nd edition) – Frackowiak, R.S.J. et al. (2004)
Offers insights into neuroimaging studies supporting localisation and distributed networks.
"Plasticity in the Human Brain: The Human Brain's Capacity for Change" – Pascual-Leone, A. (2005)
Focuses on neuroplasticity and how the brain reorganises itself after injury or learning new tasks.
"Neuroscience: Exploring the Brain" (5th edition) – Bear, M.F., Connors, B.W., & Paradiso, M.A. (2020)
An accessible text that covers localisation of function with an emphasis on recent research findings.
ONLINE RESOURCES
Society for Neuroscience (SfN): https://www.sfn.org/
Offers a variety of resources on brain localisation, neuroplasticity, and cognitive neuroscience.
BrainFacts.org: https://www.brainfacts.org/
Educational content focused on brain function, localisation, and plasticity.
PubMed: https://pubmed.ncbi.nlm.nih.gov/
Database of research articles on localisation, neuroplasticity, and distributed processing.